... és a bennük lejátszódó kémiai folyamatok
Esszémben a növények szerves és szervetlen anyagfelvételre képes részeinek sejtszintű felépítéséről és a sejtekben lejátszódó kémiai reakciókat szeretném részletezni és azok fontosságáról szeretnék említést tenni.
A növények nagy része az eukarióta törzsbe tartozik, mert az örökítőanyagot sejtmag veszi körül, nem pedig szabadon helyezkedik el a sejtben a többi sejtalkotó mellett, viszont érdemes megemlíteni, hogy prokarióta törzsbe tartozó növények is előfordulnak, így nem lehet éles határt húzni, hogy a növények csak eukarióta sejteket tartalmaznak. Mint minden sejt rendelkezik egy éles határvonallal, ami elválasztja az inter- és extracelluláris teret egymástól, ez a határvonal a sejtfal.
Sejtfal felépítése
A sejtfal anyaga nagy részben cellulózból áll, ami poliszacharid, vagyis egyszerű cukrok összekapcsolódásából jön létre. A cellulózt felépítő molekulák béta-glükóz egységek, amik cisz-, illetve transz térállásban kapcsolódnak, a képződött láncok pedig hidrogénkötésben kapcsolódnak egymáshoz, ezáltal kialakítva egy stabil és kémiailag ellenálló szerkezetet. Ezek a jellemzők segítik elő, hogy a sejtfal rugalmas és könnyen formálható legyen, hiszen ellent kell állni a sejtben uralkodó turgor nyomásnak, a cella növekedésekor bekövetkező változásoknak is.
Az elsődleges sejtfal citoszkeleton által rendezett cellulózszálak egymáshoz kapcsolódó rétegeiből kialakult plazmamembrán. Hidrofilitását tekintve hidrolfil, mert átjárható a víz és kisebb molekulák (ionok, szerves vegyületek, kis méretű fehérjék) számára. A cellulózrostok átmérője 10-25 nm, nagy szakítószilárdságúak, meghatározott rendszer szerint helyezkednek el a mátrixban, ami egy gélszerű anyag, hemicellulóz és pektin építi fel, ezek a Golgi- készülékben szintetizálódnak és a szekréciós vezikulumokban szállítódnak a sejtfalhoz, ahol ahol betöltik a funkciójukat.
A másodlagos sejtfal kötő-és érszövetekből épül fel, az elsődleges és a plazma membrán közötti teret elfoglalva. Itt is cellulózszálak helyezkednek el meghatározott rend szerint, viszont a rostok közötti teret polimerizált fenolvegyületek (lignin) töltik ki, ezzel növelve a sejtfal erősségét, emellett megakadályozza a gombafonalak és egyéb kórokozók mechanikai átjutását a sejtfalon és annak gombaeredetű enzimek általi lebontását. A cellulózrostok feladata nemcsak az elsődleges és másodlagos sejtfal felépítése, hanem áthatolnak a kettős rétegű plazmamembránon egy rosette elnevezésű, szintén cellulózrostok halmazából felépülő összekötő hídon és a plazmamembrán felületén meghatározott rendszer szerint elhelyezkedő kérgi mikrotubulusokba kapcsolódnak.
Az intercelluláris térben sejtszervecskék helyezkednek el, amik szintén citoszkeleton segítségével mozognak a citoplazmában. Finom mikrotubuláris tubulin és mikrofibrilláris aktin fehérjerostok alkotják a citoszkeletont, mintegy hálózatot alkotva a citroplazmában.
A sejt 90%-át a vakuólum (sejtnedvüreg) alkotja, tonoplaszt membrán veszi körül, feladata tápanyagok, anyagcseretermékek raktározása, transzportja, kiválasztása, illetve a sejt belső terének kitöltése, szerepet játszik az ozmotikus szabályozásban is.
Sejtszervecskék
A sejtmagvacska (nucleolus) egy gömbszerű altér a sejtmagon belül, itt történik a riboszómális RNS szintézise és itt találhatók a riboszómális alegységek. A sejtmagvacskát a sejtmag védi, ami specializált szerkezettel rendelkezik, mert külső és belső burok határolja el a sejt további részeitől. A burok felületén pórusok találhatók, amik a nagyobb molekulák számára biztosítanak átjárhatóságot. A sejtmagburok kapcsolódhat az endoplazmatikus retikulumhoz (ER), ez szintén egy membrán rendszer és két fajtája fordul elő a sejtben: a sima és a redős endoplazmatikus retikulum.
A sejtmagnak nemcsak védelmi, hanem irányító és szabályozó szerepe van a sejtnövekedésben- és anyagcsere folyamatokban, benne találhatóak a gének, amik örökletes tulajdonságokat hordoznak.
A sima endoplazmatikus retikulumban lipidszintézis folyamata megy végbe, a redős endoplazmatikus retikulumban pedig fehérjeszintézis. Növényi sejtekben az ER felosztható a citoplazmában elhelyezkedő és a sejtmagot körülvevő ER-ra.
Anyagcsere-folyamatokért felelős sejtszervecskék a mitokondrium és a kloroplasztisz (zöld színtest). Mitokondriumban adenozin-trifoszfát (ATP) képződik szénhidrátok, lipidek lebontásával, kettős membránréteggel rendelkezik.
Az adenozin-5-trifoszfát (ATP) a növényekben energiaforrásként fordul elő. Minden esetben ADP-ből keletkezik foszforiláció útján, ami kétféle úton mehet végbe. Az egyik az elektrontranszport-lánchoz kötött oxidatív foszforiláció, ennek során egy membránba ágyazott elektrontranszport-lánc a működése során proton koncentráció különbséget hoz létre a membrán két oldalán. A proton koncentráció különbség (az ún. proton-hajtóerő) lehetővé teszi , hogy a kiegyenlítődés felé ható proton átáramlásban rejlő energia ATP szintézisre fordítódjon. A protonok egy membránba ágyazott enzimen, az ATP-szintázon haladnak keresztül, és ennek során keletkezik ATP ADP-ből és szervetlen foszfátból.
Eukariótákban, ezen belül növényekben a mitokondriumban, a színtestekben (kloroplasztiszban) is zajlik ez a folyamat. Másik formája az úgynevezett szubsztrát-szintű foszforiláció. Ennek során a foszfát (helyesebben foszforil-) csoport egy foszforilált szerves molekuláról kerül az ADP-re. Legnagyobb részben a fotoszintézis (lásd növények és egyes baktériumok) és összetettebb szerves molekulák lebontása (oxidációja) során keletkezik, ez utóbbi folyamat minden élőlényben zajlik.
Kloroplasztisztban található vegyületek a klorofill A és klorofill B vegyületek adják a sejtszervecske zöld színét.
Fotoszintézis útján képződő monoszacharid a szőlőcukor, a folyamat lejátszódásához szükséges energiaforrás fényenergia és a szénforrás pedig a leveleken keresztül beáramló szén-dioxid. Zöld színtesten kívül más specifikus színtestek is megtalálhatók a sejtben, mint az amiloplasztisz (keményítő tárolása, zsírsavak és terpének, illetve más nyersanyagok előállítása, amelyek energiát biztosítanak egyéb nagymolekulák szintéziséhez), a rhodoplasztisz, ami piros színanyagot hordoz, a muro- vagy cianoplasztisz (ez ciánzöld színanyagot tárol, megtalálható az egyik algafajban), kromoplasztisz (további színanyagok szintézise és tárolása), gerontoplasztisz (fotoszintézisben résztvevő anyagok lebomlásának szabályozása a levél elhalásakor), leukoplastisz (színtelen, monoterpének szintéziséért felelős) és átalakulhat más színtestekké, mint pl. elaioplasztisz (növényi zsír tárolása), proteinoplasztisz (fehérjék tárolása és átalakítása), tannoszóma (tanninok, polifenolok szintézise). Peroxiszóma kataláz enzimet tartalmaz, ami az anyagcserefolyamatok során képződött hidrogén-perodixot eltávolítja és átalakítja.
Tulajdonképpen a citoplazma egy vízalapú folyadékból, ami a citoszol és a sejtszervecskékből áll.
A szomszédos sejtek közötti kapcsolatot a plazmodezmata biztosítja, amit egy vékony szálnak kell elképzelni és hídként viselkedik. A plazmamembrán külső felületén található térrész az apoplaszt, ami vízáteresztő tulajdonsággal rendelkezik, így a hidrofil molekulák könnyen oldódnak. A sejtek között egy megtalálható pektin poliszacharidban gazdag, ezáltal ragadós, középső lemezes felület összeragasztja a szomszédos sejteket.
A levél felszínét borító viaszréteg elsősorban hirdofób szénhidrogén vegyületekből épül fel, ezek tartalmazhatnak telített és telítetlen kötéseket, funkciós csoportokat, amik befolyásolják a vegyület hidrofóbicitását.
A többi élő szervezethez hasonlóan, a növényekben is kondenzációs reakciók játszódnak le nukleinsavak, fehérjék, polipeptidek szintézisekor. Viszont ez csak formális, mert termodinamikailag kedvezőtlen, ezért az a megoldás, hogy a kondenzáció egy másik úton megy végbe úgy, hogy a reakció során egy másik, nagy szabadentalpia csökkenéssel járó molekuláris átalakulás is végbemegy.
Az aminosavakat aktiválni szükséges ahhoz , hogy megtörténjen az összeépülés, ilyenkor nem szabadaminosav állapotban vannak, hanem egy tRNS molekulához vannak kapcsolva kovalensen, észterkötéssel. Az észterkötés elbomlása szabadentalpia csökkenést okoz. A reakció során egy amin végez nukleofil támadást egy karbonil szénatomon, de a kilépő csoport nem egy hidroxil, hanem egy tRNS molekula. Ezt az energiaigényes szintetikus folyamatot ATP-ben rejlő kémiai energia segítségével lehet végrehajtani.
Anyagcsere-folyamatok
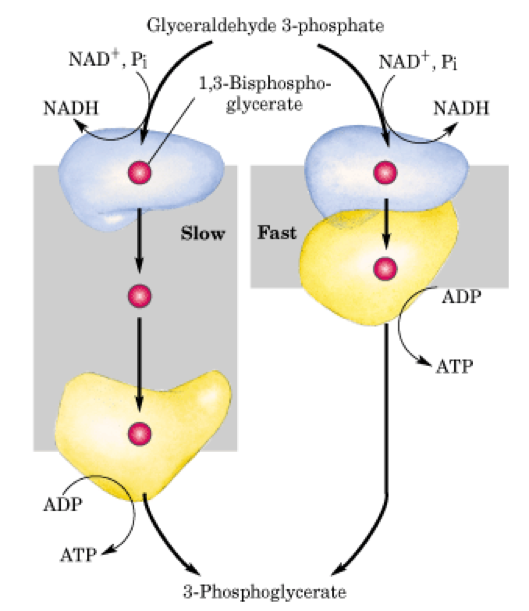
Glicenaldehid-3-foszfát átalakítása 3-foszfogliceráttá
Mindkét folyamat energiatermelő, viszont ahol 1,3 - Bifoszfoglicerát is keletkezik, egy lassú folyamat.
A citoszolban az enzimkomplexek kialakulását meghatározza a glikolitikus enzimek koncentrációja. A reakció hatékonyságát a kináz és a dehidrogenáz enzimek egymáshoz való közelsége növeli a reakció hatékonyságát és a dehidrogenáz enzim termekének eljutását a kináz enzimhez.
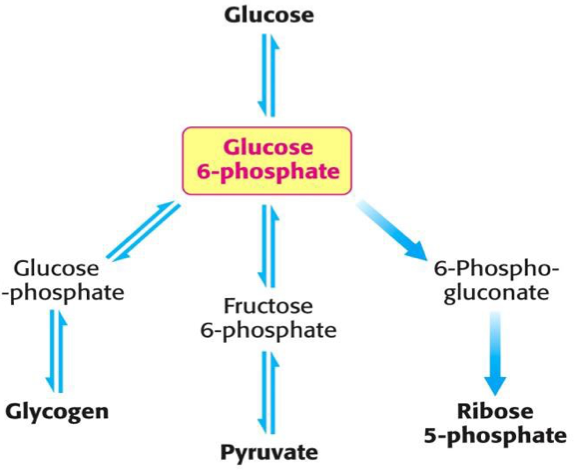
Szénhidrát anyagcsere fő útvonalai a glukóz-6-foszfátból indulnak, az egyes útvonalak az organizmus homeosztázisa érdekében többféle módon szabályozottak.
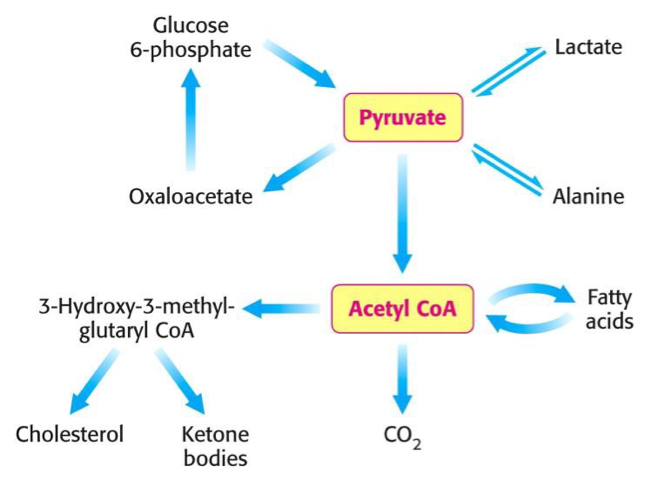
Az anyagcsere-folyamatokban képződő vegyületek közül a két legfontosabb és könnyen átalakítható vegyületek a piruvát (piroszőlősav deprotonált alakja) és az Acetil-koenzim A.
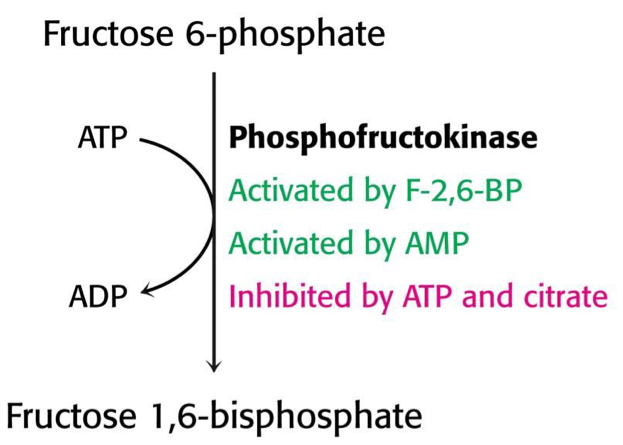
A glikolízis sebességét elsősorban a foszfofruktokináz enzim aktivitása határozza meg alloszterikus aktivátorok és inhibítorok segítségével. A fruktóz 2,6-biszfoszfát regulátor metabolit keletkezését pedig a foszforuktokináz-II enzim kovalens módosítása szabályozza.
Zsírsav szintézis és lebontás szabályozása.
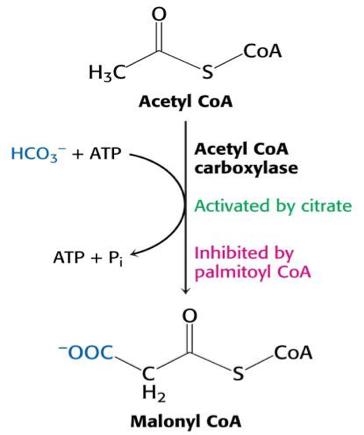
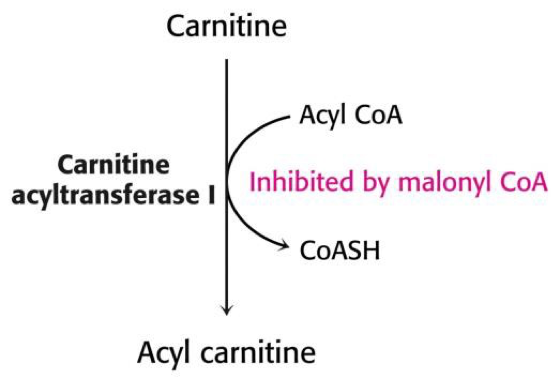
A sejtekben lejátszódó anyagcsere-folyamatok nem minden esetben tudnak az “általános” úton lejátszódni, mert egy élő rendszerről beszélünk, ahol folyamatos változások mennek végbe, amik hatással vannak egymásra. Figyelembe véve a termodinamikai és kinetikai törvényszerűségeket.